Nature encourages no looseness, pardons no errors
|

|
10 Oct 2010
|
In this lesson, we wish to ask:
- How did observations in nature lead to the
formulation of the theory of evolution?
- What are the main points of Darwin's theory of
evolution?
- How does the process of natural selection work?
- What evidence do we have for local adaptation?
- How can natural selection affect the frequency of
traits over successive generations?
The (R)Evolution of Theory
The theory of evolution is one of
the great intellectual revolutions of human history, drastically changing our
perception of the world and of our place in it. Charles Darwin put forth a
coherent theory of evolution and amassed a great body of evidence in support
of this theory. In Darwin's time, most scientists fully believed that each
organism and each adaptation was the work of the creator. Linneaus
established the system of biological classification that we use today, and
did so in the spirit of cataloguing God's creations.
In other words, all of the similarities and dissimilarities among groups of organisms that are the result of the branching process creating the great tree of life (see Figure 1), were viewed by early 19th century philosophers and scientists as a consequence of omnipotent design.
|
Figure 1: A phylogenetic "tree of life"
constructed by computer analysis of cyochrome c molecules in the organisms
shown; there are as many different trees of life as there are methods of
analysis for constructing them. |
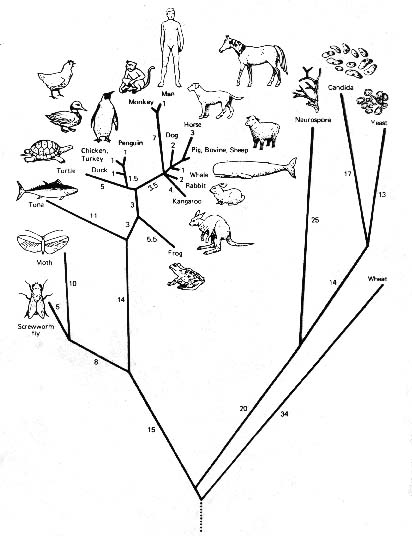
However, by the 19th Century, a number of natural historians were beginning to think of evolutionary change as an explanation for patterns observed in nature. The following ideas were part of the intellectual climate of Darwin's time.
- No one knew how old the earth was, but geologists
were beginning to make estimates that the earth was considerably older
than explained by biblical creation. Geologists were learning more about
strata, or layers formed by successive periods of the deposition
of sediments. This suggested a time sequence, with younger strata
overlying older strata.
- A concept called uniformitarianism,
due largely to the influential geologist Charles Lyell, undertook to
decipher earth history under the working hypothesis that present
conditions and processes are the key to the past, by investigating
ongoing, observable processes such as erosion and the deposition of
sediments.
- Discoveries of fossils were accumulating during the
18th and 19th centuries. At first naturalists thought they were finding
remains of unknown but still living species. As fossil finds continued,
however, it became apparent that nothing like giant dinosaurs was known
from anywhere on the planet. Furthermore, as early as 1800, Cuvier
pointed out that the deeper the strata, the less similar fossils were to
existing species.
- Similarities among groups of organisms were considered
evidence of relatedness, which in turn suggested evolutionary change.
Darwin's intellectual predecessors accepted the idea of evolutionary
relationships among organisms, but they could not provide a satisfactory
explanation for how evolution occurred.
- Lamarck is the most famous of these. In 1801, he
proposed organic evolution as the explanation for the physical
similarity among groups of organisms, and proposed a mechanism for
adaptive change based on the inheritance of acquired characteristics. He
wrote of the giraffe:
"We know that this animal, the tallest of mammals, dwells in the interior of Africa, in places where the soil, almost always arid and without herbage, obliges it to browse on trees and to strain itself continuously to reach them. This habit sustained for long, has had the result in all members of its race that the forelegs have grown longer than the hind legs and that its neck has become so stretched, that the giraffe, without standing on its hind legs, lifts its head to a height of six meters."
In essence, this says that the necks of Giraffes became long as a result of continually stretching to reach high foliage. Larmarck was incorrect in the hypothesized mechanism, of course, but his example makes clear that naturalists were thinking about the possibility of evolutionary change in the early 1800's.
- Darwin was influenced by observations made during his
youthful voyage as naturalist on the survey ship Beagle. On the
Galapagos Islands he noticed the slight variations that made tortoises from
different islands recognizably distinct. He also observed a whole array
of unique finches, the famous "Darwin's finches," that
exhibited slight differences from island to island. In addition, they
all appeared to resemble, but differ from, the common finch on the
mainland of Ecuador, 600 miles to the east. Patterns in the distribution
and similarity of organisms had an important influence of Darwin's
thinking. The picture at the top of this page is of Darwin's own
sketches of finches in his Journal of Researches.
- In 1859, Darwin published his famous On the Origin
of Species by Means of Natural Selection, a tome of over 500 pages
that marshalled extensive evidence for his theory. Publication of the
book caused a furor - every copy of the book was sold the day that it
was released. Members of the religious community, as well as some
scientific peers, were outraged by Darwin's ideas and protested. Most
scientists, however, recognized the power of Darwin's arguments. Today,
school boards still debate the validity and suitability of Darwin's
theory in science curricula, and a whole body of debate has grown up
around the controversy (see the WWW site Talk.Origins for an ongoing
dialogue). We do not have time to cover all of Darwin's evidence and
arguments, but we can examine the core ideas. What does this theory
of evolution say?
Darwin’s theory of evolution entails the following fundamental ideas. The first three ideas were already under discussion among earlier and contemporaneous naturalists working on the “species problem” as Darwin began his research. Darwin’s original contributions were the mechanism of natural selection and copious amounts of evidence for evolutionary change from many sources. He also provided thoughtful explanations of the consequences of evolution for our understanding of the history of life and modern biological diversity.
-
Species (populations of interbreeding organisms) change over time and space. The representatives of species living today differ from those that lived in the recent past, and populations in different geographic regions today differ slightly in form or behavior. These differences extend into the fossil record, which provides ample support for this claim.
-
All organisms share common ancestors with other organisms. Over time, populations may divide into different species, which share a common ancestral population. Far enough back in time, any pair of organisms shares a common ancestor. For example, humans shared a common ancestor with chimpanzees about eight million years ago, with whales about 60 million years ago, and with kangaroos over 100 million years ago. Shared ancestry explains the similarities of organisms that are classified together: their similarities reflect the inheritance of traits from a common ancestor.
-
Evolutionary change is gradual and slow in Darwin’s view. This claim was supported by the long episodes of gradual change in organisms in the fossil record and the fact that no naturalist had observed the sudden appearance of a new species in Darwin’s time. Since then, biologists and paleontologists have documented a broad spectrum of slow to rapid rates of evolutionary change within lineages.
The primary mechanism of change over time is natural selection, elaborated below. This mechanism causes changes in the properties (traits) of organisms within lineages from generation to generation.
The Process of Natural Selection
Darwin’s process of natural selection has four components.
- Variation. Organisms (within populations) exhibit individual variation in appearance and behavior. These variations may involve body size, hair color, facial markings, voice properties, or number of offspring. On the other hand, some traits show little to no variation among individuals—for example, number of eyes in vertebrates.
- Inheritance. Some traits are consistently passed on from parent to offspring. Such traits are heritable, whereas other traits are strongly influenced by environmental conditions and show weak heritability.
- High rate of population growth. Most populations have more offspring each year than local resources can support leading to a struggle for resources. Each generation experiences substantial mortality.
- Differential survival and reproduction. Individuals possessing traits well suited for the struggle for local resources will contribute more offspring to the next generation.
From one generation to the next, the struggle for resources (what Darwin called the “struggle for existence”) will favor individuals with some variations over others and thereby change the frequency of traits within the population. This process is natural selection. The traits that confer an advantage to those individuals who leave more offspring are called adaptations.
In order for natural selection to operate on a trait, the trait must possess heritable variation and must confer an advantage in the competition for resources. If one of these requirements does not occur, then the trait does not experience natural selection. (We now know that such traits may change by other evolutionary mechanisms that have been discovered since Darwin’s time.)
Natural selection operates by comparative advantage, not an absolute standard of design. “…as natural selection acts by competition for resources, it adapts the inhabitants of each country only in relation to the degree of perfection of their associates” (Charles Darwin, On the Origin of Species, 1859).
During the twentieth century, genetics was integrated with Darwin’s mechanism, allowing us to evaluate natural selection as the differential survival and reproduction of genotypes, corresponding to particular phenotypes. Natural selection can only work on existing variation within a population. Such variations arise by mutation, a change in some part of the genetic code for a trait. Mutations arise by chance and without foresight for the potential advantage or disadvantage of the mutation. In other words, variations do not arise because they are needed.
Evidence
of Natural Selection
Let's
look at an example to help make natural selection clear.
Industrial melanism is a phenomenon that affected over 70 species of moths in England. It has been best studied in the peppered moth, Biston betularia. Prior to 1800, the typical moth of the species had a light pattern (see Figure 2). Dark colored or melanic moths were rare and were therefore collectors' items.
|
|

During
the Industrial Revolution, soot and other industrial wastes darkened tree
trunks and killed off lichens. The light-colored morph of the moth became
rare and the dark morph became abundant. In 1819, the first melanic morph was
seen; by 1886, it was far more common -- illustrating rapid evolutionary
change. 
Eventually light morphs were common in only a few locales, far from industrial areas. The cause of this change was thought to be selective predation by birds, which favored camouflage coloration in the moth.
In the 1950's, the biologist Kettlewell did release-recapture experiments using both morphs. A brief summary of his results are shown below. By observing bird predation from blinds, he could confirm that conspicuousness of moth greatly influenced the chance it would be eaten.
Recapture Success
|
light moth |
dark moth |
|
|
non-industrial woods |
14.6 % |
4.7 % |
|
industrial woods |
13 % |
27.5 % |
Local Adaptation - More Examples
So
far in today's lecture we have emphasized that natural selection is the
cornerstone of evolutionary theory. It provides the mechanism for adaptive
change. Any change in the environment (such as a change in the background
color of the tree trunk that you roost on) is likely to lead to local
adaptation. Any widespread population is likely to experience different
environmental conditions in different parts of its range. As a consequence it
will soon consist of a number of sub-populations that differ slightly, or
even considerably.
The following are examples that illustrate the adaptation of populations to local conditions.
- The rat snake, Elaphe obsoleta, has
recognizably different populations in different locales of eastern
North America (see Figure 3). Whether these should be called
geographic "races" or subspecies is debatable. These
populations all comprise one species, because mating can occur between
adjacent populations, causing the species to share a common gene pool (see
the lecture on speciation).
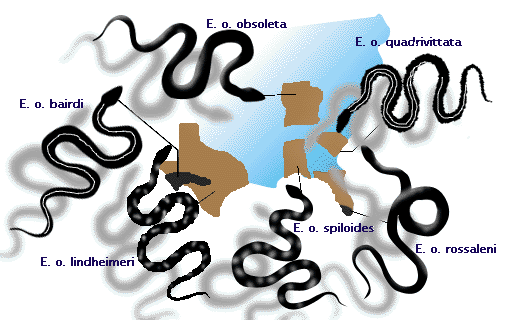
Figure 3: Subspecies of the rat snake Elaphe obsoleta, which interbreed where their ranges meet.
- Galapagos finches are the famous example from
Darwin's voyage. Each island of the Galapagos that Darwin visited had
its own kind of finch (14 in all), found nowhere else in the world.
Some had beaks adapted for eating large seeds, others for small seeds,
some had parrot-like beaks for feeding on buds and fruits, and some had
slender beaks for feeding on small insects (see Figure 4). One
used a thorn to probe for insect larvae in wood, like some woodpeckers
do. (Six were ground-dwellers, and eight were tree finches.) (This
diversification into different ecological roles, or niches, is
thought to be necessary to permit the coexistence of multiple species,
a topic we will examined in a later lecture.) To Darwin, it appeared
that each was slightly modified from an original colonist, probably the
finch on the mainland of South America, some 600 miles to the east. It
is probable that adaptive radiation led to the formation of so many
species because other birds were few or absent, leaving empty niches to
fill; and because the numerous islands of the Galapagos provided ample
opportunity for geographic isolation.
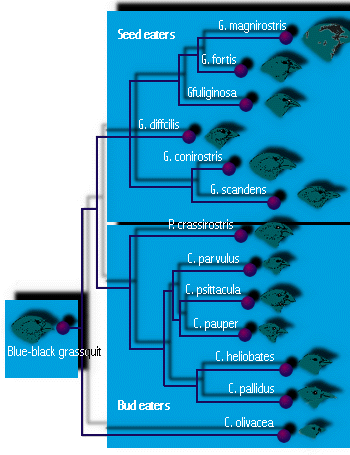
Figure 4
Stabilizing, Directional, and
Diversifying Selection
Finally,
we will look at a statistical way of thinking about selection. Suppose that
each population can be portrayed as a frequency distribution for some trait
-- beak size, for instance. Note again that variation in a trait is the
critical raw material for evolution to occur.
What will the frequency distribution look like in the next generation?
|
|
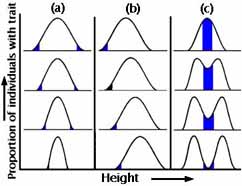
First, the proportion of individuals with each value of the trait (size of beak, or body weight) might be exactly the same. Second, there may be directional change in just one direction. Third (and with such rarity that its existence is debatable), there might be simultaneous change in both directions (e.g. both larger and smaller beaks are favored, at the expense of those of intermediate size). Figures 5a-c capture these three major categories of natural selection.
|
|
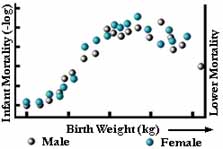
Under stabilizing selection, extreme varieties from both ends of the frequency distribution are eliminated. The frequency distribution looks exactly as it did in the generation before (see Figure 5a). Probably this is the most common form of natural selection, and we often mistake it for no selection. A real-life example is that of birth weight of human babies (see Figure 6).
Under directional selection, individuals at one end of the distribution of beak sizes do especially well, and so the frequency distribution of the trait in the subsequent generation is shifted from where it was in the parental generation (see Figure 5b). This is what we usually think of as natural selection. Industrial melanism was such an example.
|
|

The fossil lineage of the horse provides a remarkable demonstration of directional succession. The full lineage is quite complicated and is not just a simple line from the tiny dawn horse Hyracotherium of the early Eocene, to today's familiar Equus. Overall, though, the horse has evolved from a small-bodied ancestor built for moving through woodlands and thickets to its long- legged descendent built for speed on the open grassland. This evolution has involved well- documented changes in teeth, leg length, and toe structure (see Figure 7).
Under diversifying (disruptive) selection, both extremes are favored at the expense of intermediate varieties (see Figure 5c). This is uncommon, but of theoretical interest because it suggests a mechanism for species formation without geographic isolation (see the lecture on speciation).
Summary
Darwin's
theory of evolution fundamentally changed the direction of future scientific
thought, though it was built on a growing body of thought that began to
question prior ideas about the natural world.
The core of Darwin's theory is natural selection, a process that occurs over successive generations and is defined as the differential reproduction of genotypes.
Natural selection requires heritable variation in a given trait, and differential survival and reproduction associated with possession of that trait.
Examples of natural selection are well-documented, both by observation and through the fossil record.
Selection
acts on the frequency of traits, and can take the form of stabilizing,
directional, or diversifying selection.
Suggested Readings
o Darwin, C. 1959. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life. London: J. Murray.
o Futuyma, D.J. 1986. Evolutionary Biology. Sunderland, Mass: Sinauer Associates, Inc.
o Dawkins, R. 1989. The Selfish Gene. Oxford: Oxford University Press.
Copyright Regents of the University of Michigan unless noted otherwise.